目前,我主要围绕B淋巴细胞发育展开相关研究。广泛来说,B淋巴细胞通过产生针对抗原的高亲和力抗体来赋予有效和持久的适应性免疫,是体液免疫反应的重要组成部分,并在免疫记忆中起着核心作用 (Weniger et al., 2025)。在本报告中,我将围绕B淋巴细胞在不同物种中的进化情况和B淋巴细胞在机体内的发育分化过程两方面展开讨论。
B细胞在不同物种中的进化
B淋巴细胞的起源可以追溯到5亿多年前开始的有颌脊椎动物适应性免疫的进化,在脊椎动物的适应性免疫进化过程中,出现了两种类型的适应性免疫策略:1)无颌物种中的基于可变淋巴细胞受体(variable lymphocyte receptors,VLRs)的系统,由VLRA和VLRB淋巴细胞介导,2)有颌物种中的基于B细胞受体(B cell receptor,BCR)和T细胞受体(T cell receptor,TCR)的系统 (Parra et al., 2013)。适应性免疫的重要标志是淋巴细胞受体/免疫球蛋白(immunoglobulins,Igs)的巨大变异性,这使它们能够识别几乎任何抗原。在淋巴细胞的发育过程中,基因片段连接多样性、体细胞超突变(somatic hypermutation,SHM)和类别转换重组(class switch recombination,CSR)等不同机制会产生这种变异性,在Ig结构中产生多样性并大大扩增了抗体库。
软骨鱼是现存最古老的有颌脊椎动物,具有胸腺和脾脏作为淋巴器官,但缺乏骨髓和淋巴结结构,在它们体内鉴定出基于 BCR/TCR 的适应性免疫的基本分子,但未发现CSR的存在,IgM是其血清中的主要抗体 (Dooley and Flajnik, 2006)。硬骨鱼的主要淋巴组织是胸腺和肾脏,次级淋巴组织为脾脏、肠道相关淋巴组织(gut-associated lymphoid tissue,GALT),硬骨鱼没有生发中心(germinal center,GC)或有组织的淋巴结构(如淋巴结和Peyer结等) (Salinas et al., 2011)。两栖动物和爬行动物中,观察到了CSR的发生,并进一步在爬行动物中观察到了SHM的证据,成年两栖动物和爬行动物的免疫系统在许多情况下与哺乳动物相似,但没有观察到与GC、淋巴结、Peyer结等免疫结构(Zimmerman et al., 2010)。
从有颌鱼到鸟类和哺乳动物的进化过程中,B细胞Igs的多样性、特异性和亲和力增加,使得针对几乎所有病原体的适应性免疫反应逐渐增强,这种进化趋势可能至少部分是由地球上温血脊椎动物的出现推动的,微生物在较温暖的宿主中更快生长和突变的能力可能是一种选择性力量 (Hsu, 1998)。在高等脊椎动物中,进一步出现了适应性免疫高度专业化的B细胞亚群(B2细胞),其中,在灵长类和啮齿类动物中,形成了完整的在骨髓的免疫细胞发生和在次级淋巴组织的适应性免疫反应,从而实现在一生中不断补充的B细胞库 (Parra et al., 2013)。接下来,将重点介绍人类和小鼠的B细胞发育分化过程。
B细胞的发育分化
概括来说,哺乳动物B细胞发育包括从原代淋巴组织(例如,人类胎儿肝脏和胎儿/成人骨髓)开始的连续阶段,随后在次级淋巴组织(例如,人类淋巴结和脾脏)中实现功能成熟,并最终通过终末分化的浆细胞产生抗体 (LeBien and Tedder, 2008)。
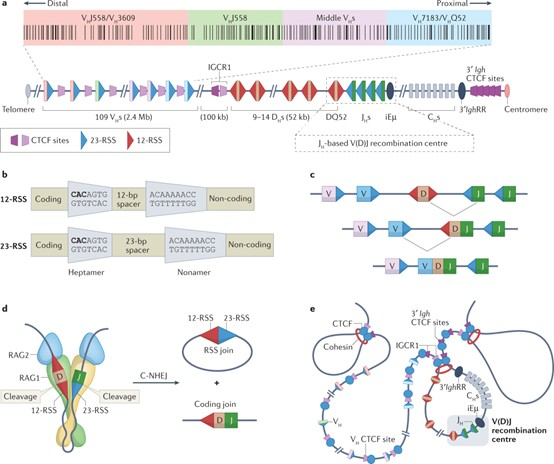
B细胞由B细胞谱系的祖细胞(pro-B细胞)在骨髓中产生的,骨髓中的多能造血干细胞产生淋巴细胞前体,从而启动不可逆的分化程序,pro-B细胞发育的特征是RAG1/RAG2参与介导的Igs的重(heavy,H)链和L(light,L)链位点的有序重排,每个pro-B细胞都经历IGH基因座的不同可变区(V)、多样区(D) 和连接区(J)基因片段的独立重排和组装,从而在每个B细胞中产生一个可变外显子,该外显子在免疫球蛋白恒定区(C)的外显子上游形成独特的序列,并驱动H链蛋白的表达(pre-B细胞)。感知到这个成熟步骤,pre-B细胞开始IGL的基因重组,这涉及将V元件组装成两个L链基因座κ或λ之一的元件,当B细胞表达L链蛋白时,它与先前重排的H链配对,在细胞表面表达为膜免疫球蛋白(immature B细胞) (Nemazee, 2017)。
接下来,immature B细胞进入次级淋巴组织的滤泡结构中,在那里它们受到T 细胞依赖性的抗原刺激并做出反应,发生增殖,分化为短寿命滤泡外浆细胞分泌抗体,或开启GC反应。GC反应的特征是B细胞的克隆扩增、IgH 基因座的CSR发生、VH基因的SHM以及通过亲和成熟增加BCR对其独特抗原表位的亲和力的选择 (Weniger et al., 2025)。其中,CSR和SHM由激活诱导的胞苷脱氨酶(Activation-induced cytidine deaminase,AID)介导 (Muramatsu et al., 2000)。在GC中,通过SHM和阳性选择产生更高亲和力的B细胞克隆, 并形成了体液免疫的记忆B细胞,这些记忆B细胞在抗原攻击后持续存在,在下一次发生对应抗原刺激反应期间迅速扩增,并可终末分化为分泌抗体的浆细胞 (LeBien and Tedder, 2008)。